

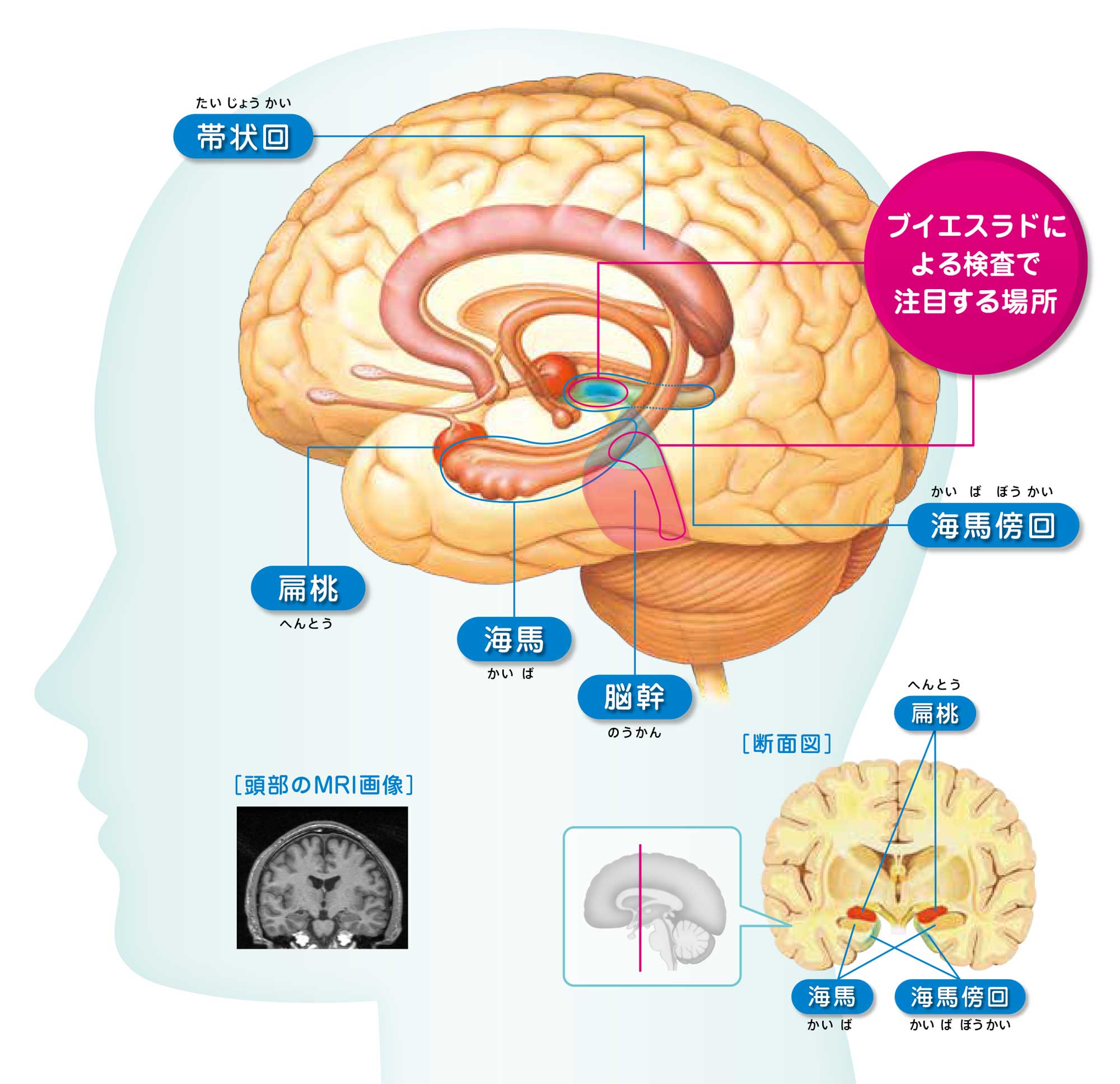
記憶に関係する海馬について
タツノオトシゴに似た形なのでこのように呼ばれます。また、ギリシャ神話に登場する海神ポセイドン(ネプチューン)がまたがる海馬(4頭立ての馬車を引く架空の動物)の尾に形が似ていることから、ルネサンス後期のイタリアで活躍したボロ-ニャ大学の解剖学者アランティオ(Arantio)が、1587年にこの脳部位を「海馬(Hippocampus)と名付けました。
.png)
https://itamikenkiyu.jimdo.com/から引用しました。
ギリシャ語でHippoは「馬」を、Kamposは「海獣」を意味し海馬は雄羊の角に似ていることから別名として「アンモン角(Ammon’s horn)」と呼ばれることもあるそうです(エジプト神アンモンは羊の角を持っている)。
小指ほどの海馬は 哺乳類の中枢神経系のなかで、もっとも詳しく研究されています。なぜでしょうか。
2つの理由があるようです。
1)解剖学的にも組織学的にも、ひと目でそれと分かる明確な構造をもっているためです。神経細胞の細胞体と、その神経網のゾーンが層状に並んでいます。
海馬体は、
細胞層の場所 細胞層 細胞層の上部 細胞層の下部
組織分類 単層 低細胞密度の層 無細胞層
部位 各層の名称
歯状回(dentate gyrus) 果粒細胞層 分子層 多形細胞層
海馬 錐体細胞層 (省略) (省略)
海馬支脚(subiculum) (省略) (省略) (省略)
前海馬支脚(presubiculum) 複数の層
傍海馬支脚(parasubiculum) 複数の層
嗅内皮質(entorhinal cortex) 複数の層
海馬の錐体細胞層は細胞の大きさと形から、さらにCA1、CA2、CA3と3つに分類されます。CAは、フランスの解剖学者ガレンジョ(Garengeot)が1742年に「アンモン角(cornu ammonis)」と名づけたことに由来しています。
海馬は二つの主要な部位にはっきりと分けることができます。歯状回に近い側の、大きな細胞がある領域と、遠い側であるより小さな細胞のある領域です。Ramon y Cajal(1911)は、これら二つの領域をそれぞれ「regio inferior」「regio superior」と名づけました。CA3野とCA2野がregio inferiorに相当し、CA1野がregio superiorに相当します。
CA3野とCA1野に存在する錐体細胞は、細胞の大きさだけでなく、神経回路網も異なり、CA3野の錐体細胞は歯状回から苔状線維の入力を受けていますが、CA2野とCA1野の錐体細胞は受けていません。
とりわけ歯状回と海馬にみられる単純な層構造は、神経解剖学や電気生理学の研究対象になっているようです。現在知られている中枢神経系のシナプス伝達に関する知見の多くは海馬体の研究に基づいたものです。
2)1950年代前半から海馬がある種の記憶や学習に基本的な役割を演じていることが認知されたからです。1957年に、HMというイニシャルをもつ、てんかん患者の治療で、新しい情報を長期記憶として、とどめることができないことが判り、海馬が注目されました。
また、てんかん発作の感受性の点で、海馬はもっとも発作閾値が低い脳部位です。ほとんどのてんかん患者は海馬が起始となった発作を示し、薬で治療するのが困難なようです。海馬体の一部分、とりわけ嗅内皮質は、アルツハイマー病でもっとも先に病変が現れ、虚血や無酸素状態に高い脆弱性を示します。海馬は、解剖学的および機能的なその構造から、ほかの大脳皮質システムのモデルになっています。
さて、海馬は記憶の形成にかかわっていますが、ここで、おさらいです。
記憶には、種類がありました。大きく分けて、陳述記憶と非陳述記憶です。
陳述記憶は、知識や過去の体験などを言葉によって、再生できます。まずは、記憶し再生できる時間の長さで短期記憶と長期記憶に分けます。
短期記憶は、電話番号を覚え、電話をかけた後、忘れるような記憶です。
長期記憶は、記憶する事柄によって、出来事記憶と意味記憶に分けられます。
出来事記憶は、個人の日々の経験や出来事についてです。意味記憶は言葉の意味や固有名詞などの知識にかかわる記憶です。
非陳述記憶は、手続き記憶です。つまり、自転車乗りや楽器の演奏などです。自然な流れで記憶が再生され、あまり考えずに、動作行えます。
海馬での大きな役割は、出来事記憶に関係するものです。
再度、おさらいです。両側の海馬を含む側頭葉内側面を手術で除去されたHMという患者は、手術後の日常の出来事の記憶がほとんどなくなりました。診察直後の記憶もなくなり、手術以前の数年間の出来事も覚えていませんでした。一方、子供のころのできgとはよく覚えていました。また、知識は正常で、手続き記憶に問題はありませんでした。
両側の海馬を含んだ損傷で、健忘が生じる部位としては、以下のものがあり、出来事記憶にかかわるものでした。
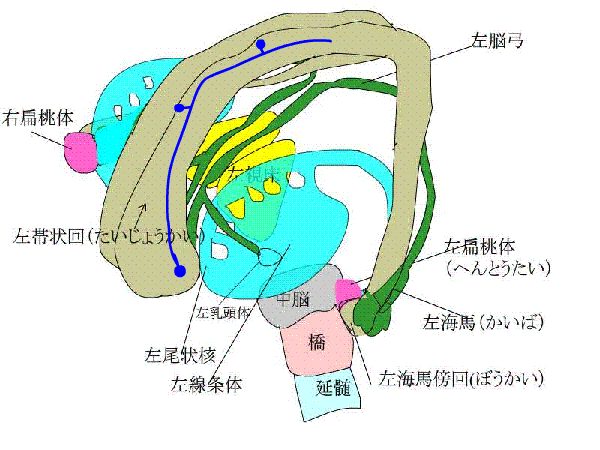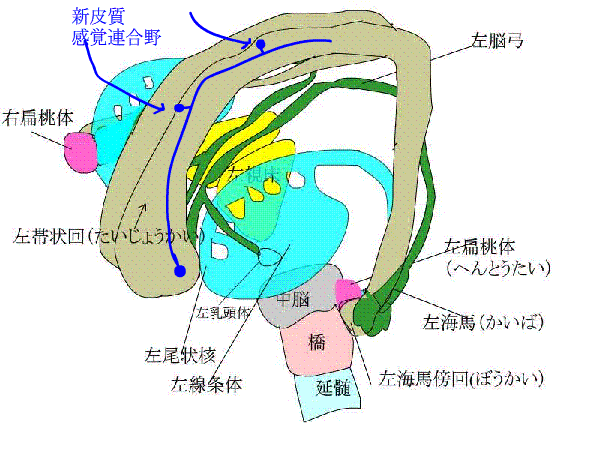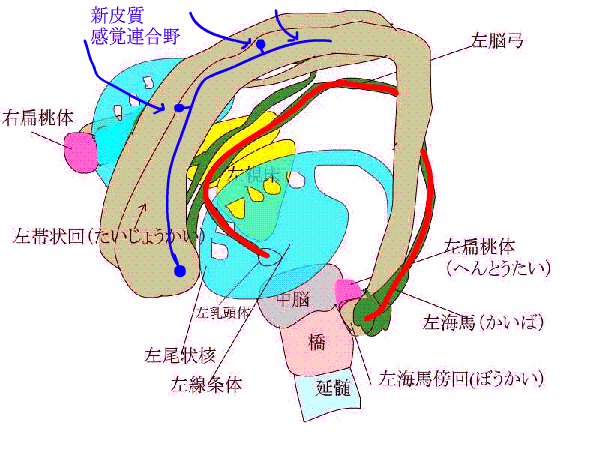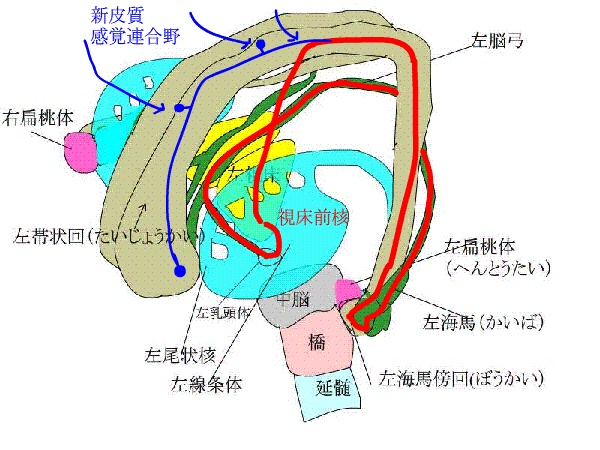
視床背内側核、視床前核、乳頭体、脳弓、大脳基底部があるようです。整理すると、海馬ー脳弓ー乳頭体ー視床前核ー帯状回ー帯状束ー海馬傍回ー海馬を結ぶ回路が存在します。
これをPapez(ペーペズ)の回路といいます。出来事記憶にかかわるものです。
記憶するには、まず、外からの情報が必要でしょう。視覚、聴覚、体性感覚(嗅覚は除いて)、味覚と痛覚の
情報が、すべて視床を経由して後頭葉、側頭葉などに集まります。
次に、これらの感覚情報の一部は、海馬に集まります。
下のアニメで、青い矢印は、新皮質と感覚連合野がある大脳皮質から、信号が辺縁系にはいることを表しています。
赤い線は、ペーペズ(Papez)の回路といって、海馬ー脳弓ー乳頭体ー視床前核ー帯状回ー帯状束ー海馬傍回ー海馬をぐるぐる回る神経ネットワークを表しています。
また、種々の連合野や辺縁皮質からーーーーー>海馬傍回に神経が伸び、
海馬傍回から---->ほとんど全ての大脳皮質に神経がはりめぐっています。
出来事記憶の中枢は、内側側頭葉,間脳(視床・乳頭体),前脳基底部でしょうか。
Papez回路では、ぐるぐると情報が回っていますが、
各個人の大脳皮質が必要と判断したとき、必要な記憶は大脳皮質連合野に格納されます。
そうでなければ、また、海馬に戻り繰り返します。
そのうち、格納する必要がない場合、情報は消えていくようです。
海馬の中をさらに詳しく見ると、
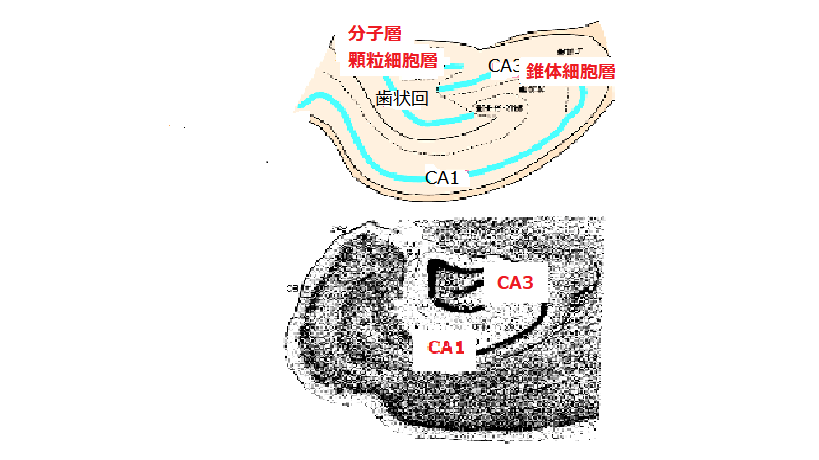
海馬の基礎知識 から引用し、改変しました。 http://gaya.jp/research/hippocampus.htm
この海馬内で、様々な神経回路があります。以下に、示しました。この図は、日本医事新報社の人体の正常構造と機能 VII神経系(I)46ページから、引用しました。本図は、ほとんど、原画のままでした。あしからず。
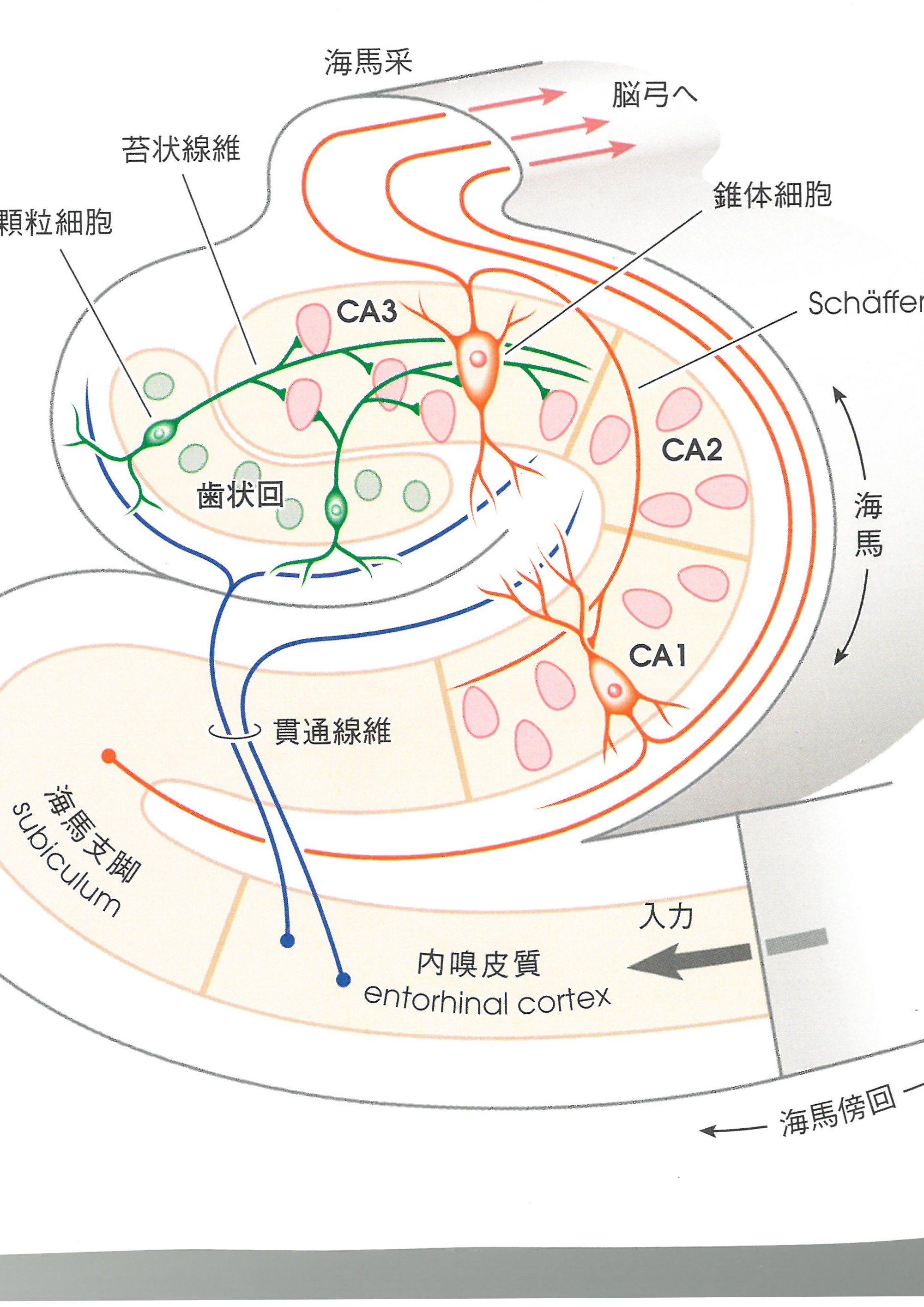
この図で、CA3領域の錐体細胞の軸索は、Schaffer側枝を出しています。
このSchaffer側枝は、CA1領域の錐体細胞に興奮性シナプスとして結合しています。このSchaffer側枝を電気刺激すると、CA1の錐体細胞に興奮性シナプス後電位が発生します。側枝を数秒間高頻度で刺激したあとでは、興奮性シナプス後電位の振幅が増大し、刺激を止めた後も、振幅の増大は、数時間以上続きます。
これが、LTP( long-term potentiation)です。これについては、グルタミン酸受容体 のコーナーで、説明しました。
LTPは、長期記憶に関連がある現象として、推測されているようです。
では、意味記憶の神経回路は何でしょうか。自分が調べる範囲では、明確な論文は見つかりませんでした。意味記憶の文字で検索すると、papezの回路がヒットします。しかし、意味記憶についての説明はありません。
判っていることは、意味記憶の中枢、つまり、意味記憶として情報が蓄えられる場所は、側頭葉のようです。
神経基盤としては 側頭葉前方部の限局性萎縮で、単語、物品、人物などの意味理解が選択的に障害されるようです。 左右差を伴い、左側頭葉前方部の萎縮では語義の障害が、右側頭葉前方部の萎縮では人物に関する記憶が障害されることが多いようです。
ここからは、井ノ口 馨先生が「記憶をあやつる」という書籍で記載された内容に沿って、ページを作成しました。ほぼ、先生の解説に基づいたものです。
場所細胞(Place cell)
この言葉は、聞きなれないですよね。今までの話は、陳述記憶(出来事記憶、意味記憶)、非陳述記憶(手続き記憶、条件反射)でした。
急に、場所細胞といわれても、ピンときません。しかし、この言葉は、1971年に、オキーフという学者が論文にしていました。
海馬に電極を刺してあるラットが、勝手に動く状態で、観察されました。すると、A地点に行った時だけ活動する神経細胞集団(a)を見つけました。ラットが、A地点から離れると、その細胞集団(a)は、活動をやめ、別の細胞集団が活動しました。この現象は、脳が常にどこかがランダムに活動しているため、不思議ではありません。
ところが、別の場所から、A地点にもどったとき、細胞集団(a)が活動したのです。それぞれ特定の場所では特定の細胞集団が活動することが判りました。
また、さらに珍しいことが判りました。先ほどのA地点に行かなければ、二度とその細胞集団は活動しないはずです。
ところが、ラットが寝ているとき、休憩しているときに、場所の特定に使われた細胞集団が、その場所を覚えたときと同じ順番で勝手に活動していることが発見されました。
これを「リプレイ」といいます。この現象は、情報を思い出すことで、記憶を固定化する、あるいは海馬から大脳皮質へと記憶をコピーするための作業かもしれません。
さらに、2011年には、以下のアニメで示すような行動がラットで見られました。
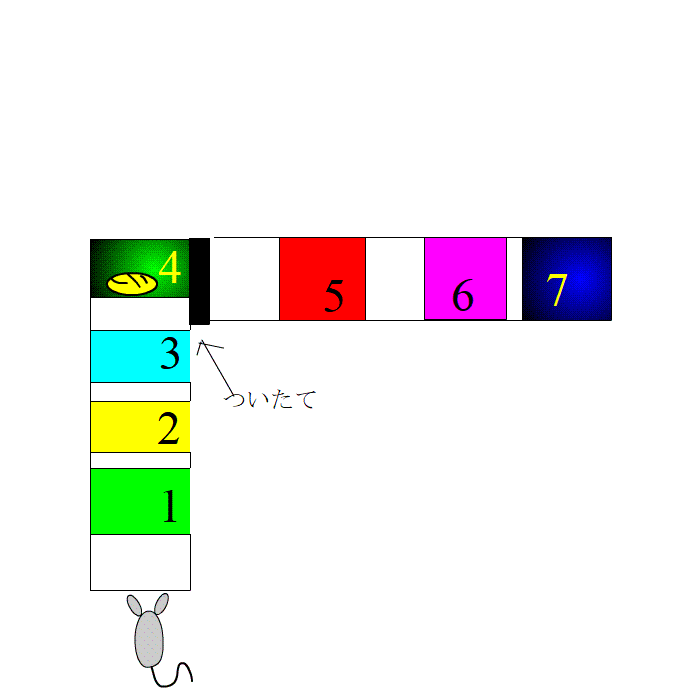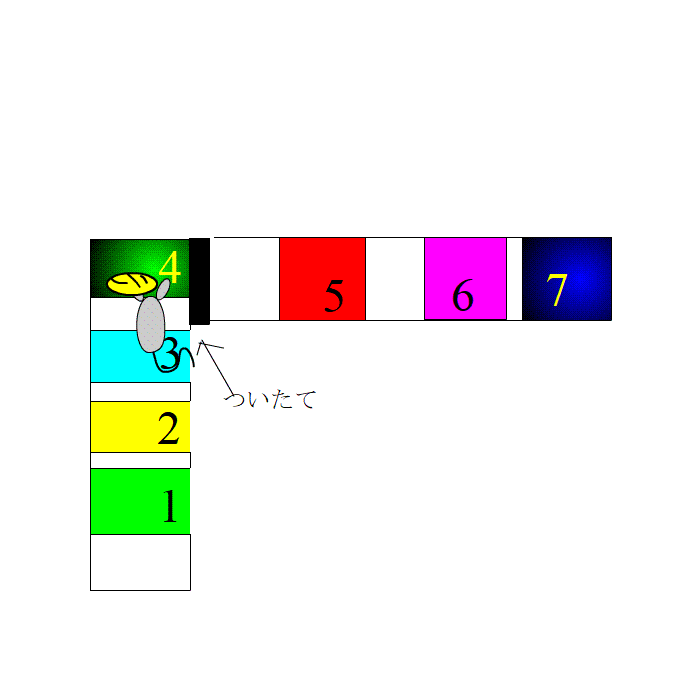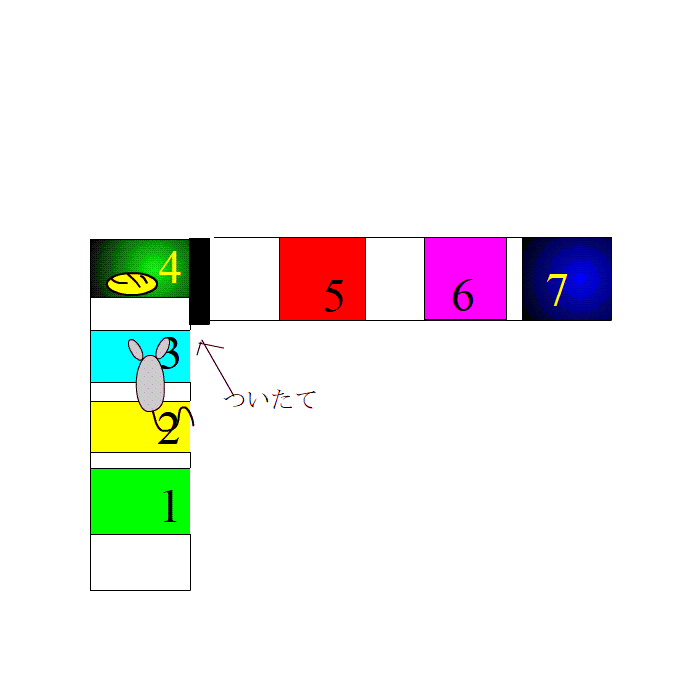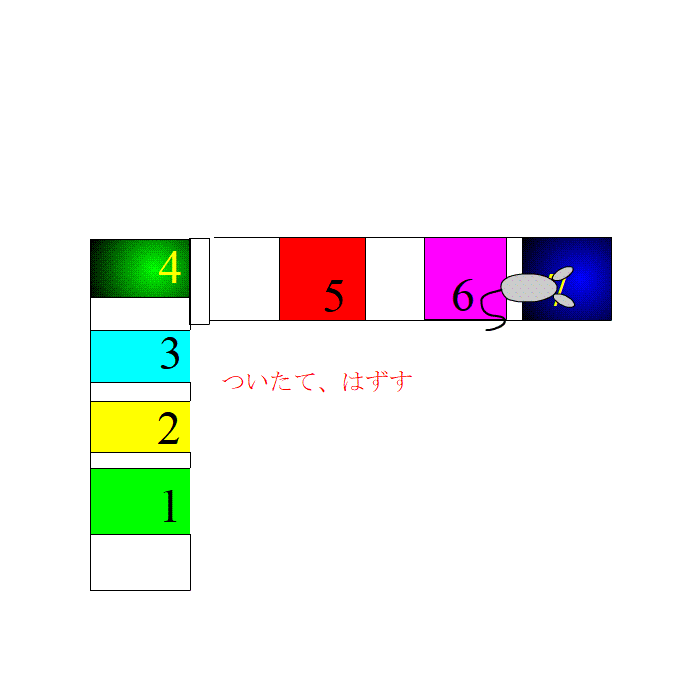
L字型の迷路です。ラットは、迷路の外にある様々な目印は見えるようになっています。
ついたての前には、餌があります。ここで、休憩ができます。
さて、実験です。ラットは、1,2,3番のプレートを走り、4番の餌場に向かいます。餌場で、えさを食べて、ゆっくりします。これを何度も繰り返します。脳内に、1,2,3とそれぞれ違う数字の場所を通るたびに、細胞集団がセットされます。
餌場でリラックスしているとき、ラットの脳内では、先ほど発火した場所細胞が発火します。つまり、リプレイの状態です。
ここで、面白いことがありました。
このとき、同じ領域の神経細胞で、リプレイに関係のない細胞の集団がランダムに発火し始めました。ノイズのようなものです。
次に、ついたてをはずしました。ラットは、新しい場所に進みました。当然、別の場所細胞が発火するわけですが、ノイズと思われていた細胞が、5,6、7と順番通りに発火しました。ラットが予知能力を持っているわけではないでしょう。
もし休憩時のランダムな自然発火が、次に来るべきじょうほうをセットするための準備を行っているサインとしたら、どうでしょう。
このように、脳細胞は、慌てて細胞集団を用意するのではなく、先に入れ物だけを用意して、いざとなったら、反応するという記憶の効率化をしているのかもしれません。
2005年には、場所細胞以外に「格子細胞」という細胞が海馬近傍の嗅内皮質に存在することが判りました。
これについては、https://www.ted.com/talks/neil_burgess_how_your_brain_tells_you_where_you_are/transcript?source=google_plusone&language=ja#t-522646 をご覧ください。
セル・アセンブリ仮説の証明について
1948年に、神経回路の可塑性(神経系に変形が起きた場合、その変形がそのまま残る性質)が記憶の本質といわれましたが、確認されずに、63年が過ぎ、2011年に証明されました。利根川 進先生によるものでした。
セル・アセンブリを証明するためには、
①学習時に活動した神経細胞の集団が、記憶の獲得や保持に本当に必要なのか。
②学習時に活動した神経細胞の集団だけを人工的に活動させたとき、その神経細胞集団に保持された記憶が本当に蘇るのか。
①は、カナダのシーナ・ジョスリンという学者が証明しました。
マウスの音恐怖条件付けという学習と記憶に一般的な手法を用いました。
実験用の箱に入れたマウスに、ブザー音を30秒ほど、聞かせます。ブザー音の最後の1~2秒間、床面に弱い電流を流して、マウスがすくみ反応が出るようにします。このマウスは、音と電気ショックを関連付けて記憶し、音を聞くだけで、すくみ反応を示します。 次に、音恐怖条件付けに重要な役割を果たす偏桃体に存在する神経細胞の約15%に、ジフテリア毒素の受容体遺伝子と同時にCREB((cAMP response element binding protein)という転写因子の遺伝子も同じ神経細胞に導入しました。CREBというたんぱく質は、たくさん作られます。 CREBをたくさん作る神経細胞は、ビックリ体験をしたときに活動しやすくなり、その結果、記憶のセル・アセンブリを形成しやすいのです。 |
この操作で、CREBとジフテリア毒素の受容体を作る神経細胞群が優先的にセル・アセンブリに取り込まれます。 ここで、マウスの脳にジフテリア毒素を流し込みました。ビックリ体験で働いた神経細胞のみがジフテリア毒素と結び付いて、そこだけ死にました。ほかの記憶は正常に働いていました。もともと、マウスには、ジフテリア毒素に結合する受容体がないため、この結果になりました。 学習するときに活動した神経細胞集団がなければ、記憶を保持したり思い出したりすることができないことが証明されました。 |
CREBの解説は、https://ja.wikipedia.org/wiki/CREB
または、https://sato-ayumi.com/ の転写因子・基本転写因子の項目を参照してください。
次に、
| 逆に、学習するときに活動した神経細胞集団を人工的に活動させ、外的な条件とは関係なく、ビックリ体験を思い出すという現象を確認することが重要でした。 |
光遺伝学という新しい研究手法を用いた利根川先生が実験を行いました。
遺伝子工学的手法を用いて、光で活性化するイオンチャネル(チャネルロドプシン)を特定の神経細胞に強制発現させることで、これらの神経細胞をピンポイントで自在に神経細胞を興奮させることができます。

記憶が出来上がったマウスの海馬に挿入しておいた光ファイバーを通してビックリ体験時に活動した神経細胞のみを活性化させました。
環境に関係なく、マウスはすくみ反応を示しました。
つまり、
| 「記憶を思い出した」状態を人工的に作り出しました。ここで、セル・アセンブリ説は証明されました。 |
私たちの記憶の一つ一つは、特定の神経細胞集団によって保持され、それが活性化すると思い出すことになります。
記憶と神経細胞の新生について
1962年、ジョセフ・アルトマンは、成獣に達したラットの海馬の細胞が増えていることに気づきました。1998年には、、人間の海馬でも神経幹細胞があり細胞が新生されていることが実証されました。
新しく生まれてくる神経細胞を殺してしまうと記憶力が低下するといわれていました。そこで、実験が行われました。
普通のマウス、
神経新生が低下するように処置したマウス
神経新生を促進させる環境で育ったマウス
これらの3種類のマウスで、利根川グループが行ったものと同じ文脈性恐怖条件付けをして、ビックリ体験の記憶が海馬依存的な記憶から非依存的記憶に変わる過程を観察しました。
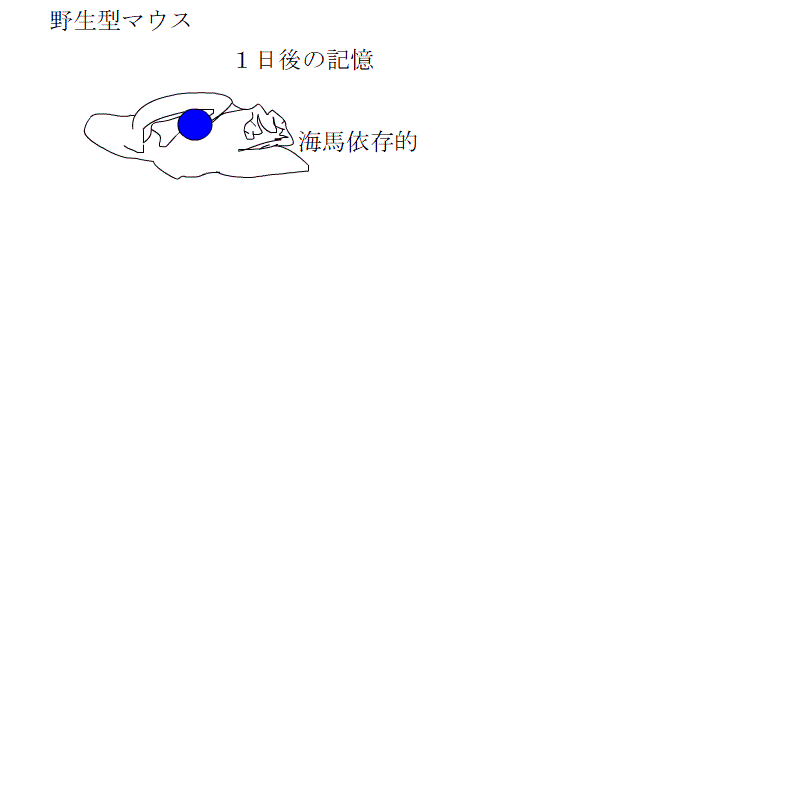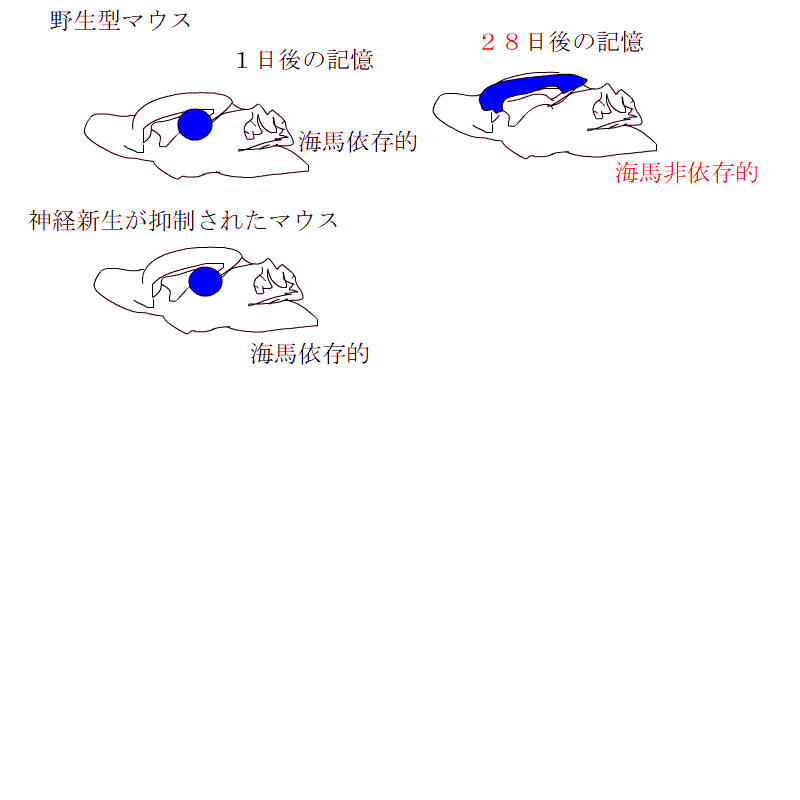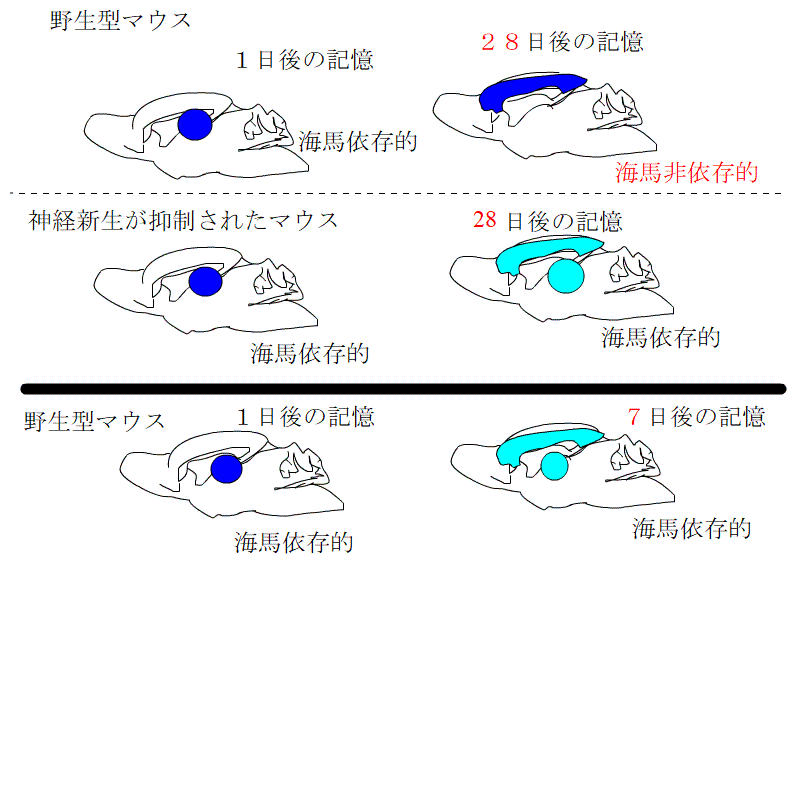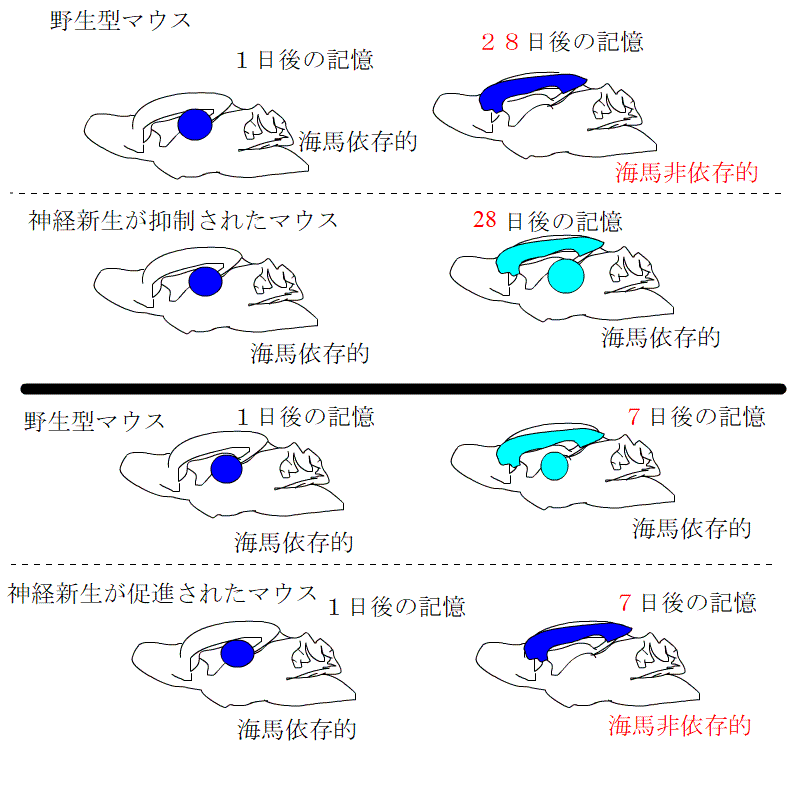
神経新生が低下したグループでは、海馬依存の期間が長くなるのに対し、促進されたグループでは、海馬依存の期間が短くなるとともに、海馬に記憶が保存される期間も短くなっていました。
つまり、長期記憶となった記憶は、一定の期間が過ぎれば、脳のほかの場所に移動させ、海馬から消し去るという作業が行われています。
| 記憶同士の関連性について |
一個の出来事には、必ず複数の情報が含まれています。つまり、記憶として保持するためには、脳の各部から送られてくる複数の情報を一個の記憶としてまとめる必要があります。
夜道を歩いていたという記憶、 ABCの細胞集団
犬がいたという記憶、 ADEの細胞集団
信号が赤に変わったという記憶があるとします。AFGの細胞集団

夜道を歩いていたことを思い出すと、同時にADEとAFGも刺激され、記憶がよみがえります。このとき、それぞれが細胞集団として独立しているため、記憶は混ざりません。このように、別々に割りつけられた記憶が関連付けられることを記憶の連合といいます。
たとえば、ブザー音を聞くとフリーズするよう恐怖条件付けをしたマウスに、ライトで光を当てつつ、同じブザーを聞かせるという実験を何度も繰り返した場合、
光だけを当てても、恐怖反応を示すようになりました。光は恐怖条件付けの条件でなかったため、これが記憶の連合になります。
この連合の仕組みを解明する仕事は、井ノ口先生が精力的に行っておられます。
神経細胞体とシナプス間で、遺伝子の発現で合成された記憶保持用たんぱく質(可塑性関連たんぱく質PRP:plasticity-relatedprotein)が関与し、シナプスタグというPRPを受け入れる仕組みで、シナプスが特異的に記憶の連合を保持するものと思います。
記憶の連合については、井ノ口先生の「記憶をあやつる」(角川選書)をお読みください。

これ以外に
記憶形成のメカニズム:分子・細胞認知学の展開 井ノ口 馨
http://www.jbsoc.or.jp/seika/wp-content/uploads/2013/05/83-02-03.pdf
新しい経験を記憶する新たな神経細胞集団を発見ー富山大ら
http://www.qlifepro.com/news/20190619/new-experience-memory-nerve-cells.htm
以上の文献も執筆されておられます。