

記憶の続き
長期増強(long-term potentiation:LTP)という現象があります。
これには、初期長期増強(early LTP)と後期長期増強(late LTP)があるようです。
海馬の内部構造は、その神経の集まり方から、歯状回、海馬台、アンモン角(CA1,CA3)に分けられますが、
その海馬歯状回のシナプスを高い周波数で刺激すると、シナプス伝達の効率が上昇し、その上昇がしばらく続きます。
この現象を長期増強(Long-Term Potentiation)となずけられました。
海馬に電極を刺し込んで、神経細胞を一秒間に数百回という高い周波数で刺激し、神経細胞を強く活性化させます。
するとシナプス電位は瞬時に大きくなり、その後、元の電位に戻るのではなく、電位が増大し持続します。
この増大した電位は、数時間から数日間も持続したのです。これが長期増強(LTP)です。
また、このLTPを引き起こすための高周波数の電気刺激をテタヌスと言います。
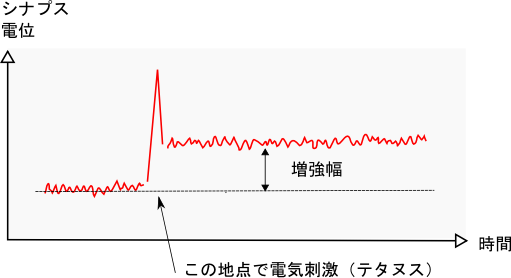
海馬にテタヌスを1回与えると、一瞬にしてシナプス電位が大きくなり、そのレベルを保持します。このことは、シナプスはテタヌスが入力されたことを記憶しているということです。実はこれが記憶のメカニズムなのです。
人が何かを記憶しているということは、刺激が海馬に電気信号として伝わり、海馬の神経細胞でLTPが起こり、電位の増大された部分が記憶になるのでしょう。
引用先居http://yorokobuya.com/index.php?%E9%95%B7%E6%9C%9F%E5%A2%97%E5%BC%B7
刺激がある一定の範囲を超えると、early LTP、late LTPの順番で起こります。
特に、late LTPには、遺伝子の発現が必要なようです。
| 身の回りで起こっている出来事すべてを認識し、記憶する 必要はないですから、受ける刺激の頻度や強さによって認識のレベルを使い分け、必要に応じてシナプス結合を強化する、つまり、記憶を作っているのでしょう。 |
一方で、逆の現象も確認されたようです。
つまり、シナプスの伝達効率が低下する長期抑圧(long -term depression :LTD)です。
長期抑圧は、主に小脳が司る運動学習に重要な役割を果たしています。
運動学習は、体で覚える学習、たとえば歩く、泳ぐなどの事です。非陳述記憶です。
非陳述記憶は、主に小脳の分担で、プルキンエ細胞という神経細胞が担っています。
プルキンエ細胞は、先端に迷路のように広がる大きな樹状突起があります。
この樹状突起には、平行繊維と登上繊維という2つの神経繊維がつながっており、
平行繊維から情報がきて、シナプスが増強されます。つまり、訓練するたびに、プルキンエ細胞への入力が大きくなります。
しかし、体が間違った動きをすると、登上繊維から「その動きはおかしいですよ、間違っています。」というエラーメッセージが、シナプスを介して、入力されます。
そのとき、間違った動きを伝達している平行繊維とつながるシナプスの伝達効率が下がり、エラーの原因になっているシナプスが神経回路から除去されるようです。
登上繊維が、「これは覚えてはだめ」と教えます。つまり、正しい動きを増強せず、間違った動きを押さえることで、伝えたくない信号を抑えるという形で学習しています。
ここで、おさらいです。
以上のような現象を理解するためには、シナプス可塑性とセル・アセンブリということの関係が重要のようです。
シナプス可塑性とは、 可塑性は、英語のplasticityの訳語で、その語源はplsticです。つまり、力を加えると形が変わり、力が除かれても、形はもとに戻りません。 つまり、変形したままの形が残る性質を表しています。神経回路の可塑性(神経系に変形が起きた場合、その変形がそのまま残る性質)が記憶の本質と結論付けました。 セル・アセンブリ(cell assembly)とは、cellは細胞、assemblyは、特定の目的のための集まりを意味します。 特定の仕事を請け負った神経細胞グループを表します。 |
わかりにくい話ですよね。つまり、
頭脳に、情報がはいり、特定の神経細胞が刺激された場合、その神経細胞とシナプスでつながる複数の神経細胞が、一つのグループ(セル・アセンブリ)を作り活動するのです。
大部分の遺伝子はそれらがコード化しているタンパク質として発現する。この過程は 2 つの流れで起こる:
- 転写 = DNA → RNA
- 翻訳 = RNA → タンパク質
概要を示す模式図

このページでは最初のステップについて述べる:
遺伝子の転写:DNA → RNA
DNA 自身の?複製?replication の時と同じように,DNA が RNA の合成のためのテンプレート ( 鋳型 ) として使われる。転写のステップ
- いくつかの?転写調節因子?transcription factors が?プロモーター?promoter 部位( たいてい転写される遺伝子の 5' 末端側 )に結合する。
- RNA ポリメラーゼ?polymerase が転写調節因子に結合する。
- これらの共同作業によって,DNA 二重らせんが局所的に解離する。
- RNA ポリメラーゼがそのうちの 1 本鎖に 3' → 5' 方向へ移動しながら作用する。
- 同時に,リボヌクレオチド三リン酸を基質に RNA が合成される。
- 塩基の対合?のルールに従って相補的な RNA を合成する。すなわち,DNA 上の C,T,G,A の各塩基が,それぞれと相補的に対合する G,A,C,U ( ウラシル ) として RNA 上に写し取られる。
- RNA 鎖は 5' 末端から 3' 末端の方向へ伸長する。
- リボヌクレオチド三リン酸が伸張中の 3' 末端に付加され,2 個のリン酸基が除去される。
- DNA 上の?転写終結?termination signal を指令する塩基配列部位で,タンパク質性の転写終結因子の介在のもとに合成を停止し,DNA より離れていく。ゲノムによっていろいろな転写終結の配列がある。
細胞に吸着したウイルスは、自らのゲノムを細胞内に送り込み、細胞がもつ転写翻訳機構を利用してゲノムを複製し増殖する。遺伝子操作により、複製および増殖能を欠損させたウイルス(増殖力欠損株)や、複製・増殖能の一部を保持したウイルスに外来遺伝子を組み込み、効率的に目的の遺伝子を細胞へ導入し発現させる能力を利用したものをウイルスベクターという。細胞への遺伝子導入効率は、エレクトロポレーションやリン酸カルシウム法などの物理化学的な導入法よりはるかに優れており、遺伝子導入が困難な生体ニューロンへの遺伝子発現実験や遺伝子治療に広く使用されている。