

グルタミン酸の受容体について
今までの話で、中枢神経系では興奮する神経の伝達物質はグルタミン酸であることが分かったと思います。
では、グルタミン酸がどのような受容体に結合するかを見てみます。
大きく分けて、2種類あります。
イオンチャネル型グルタミン酸受容体、つまり、陽イオンを透過させるもの。 もう一つは、チャネル活性はないが、グルタミン酸が結合することで、細胞内のセカンドメッセンジャー(Ca++, IP3, cAMP)の産生を調節するG蛋白共役型です。 |
このイオンチャネル型については、さらに3種類あります。
AMPA型受容体 NMDA型受容体 Kinate受容体 |
AMPA型受容体
人工アミノ酸であるAMPA(α-アミノ-3-ヒドロキシ-5-メソオキサゾール-4-プロピオン酸)を選択的に受容することから名づけられました。
これが名前の由来です。
中枢神経系に広く分布し、思考、認識、記憶や学習に大きく関与します。中枢神経系におけるグルタミン酸のシナプス興奮性伝達は、普段主にこの受容体によって行われています。
サブユニットで構成され、GluR1,GluR2, GluR3, GluR4の4つのサブユニットがわかっています。
これらが集まり4量体を形作っています。サブユニットの組み合わせについてはまだ議論の余地があり、定説はないそうです。
それぞれのGluRサブユニットには一つずつリガンドを受容する部位があり、よって全てのAMPA受容体は4つのリガンド受容部位を持つことになります。このうち、2つ以上にリガンドが結合する事でイオンチャネル部位が開くとされているようです。
NMDA型受容体
N-メチル-D-アスパラギン酸です。この物質を受容するため、名づけられました。
注)リガンド(Ligand; ライガンド)とは、生体分子と複合体を形成して生物学的な目的を果たす物質のことを指します。 タンパク質-リガンド結合では、リガンドは通常、標的タンパク質上の結合部位に結合することでシグナルを生成する分子です。
注)サブユニットとは、蛋白質の完成した1分子が,数個のポリペプチド鎖の組合せから成るとき,各1本のポリペプチド鎖を,単位分子のまとまった一部分ずつという意味で,サブユニットといいます。
AMAPAとNMDA受容体の特徴
哺乳動物の脳内では、速い(数十msec)膜電位の変化を司る分子は、イオンチャネル型グルタミン酸受容体です。また、ほとんどがカルシウム非透過性ですが、一部のAMPAとNMDA型は、カルシウムを通し、シナプス可塑性には必須です。 通常チャネルを閉じているNMDA受容体は、強い刺激で膜電位が上昇した際、チャネルを閉じていたマグネシウムがはずれ、カルシウムを含む陽イオンを通します。下のアニメ参照 |
AMPA・NMDA受容体とLTP/LTDの関係
改めて、長期増強(long term potentiation:LTP)とは何でしょうか。???
LTPという現象は、シナプスの可塑性を研究する中で、発見された出来事で、記憶そのものを表現しているものではないようです。LTPが障害されると、記憶の形成や維持が障害(特に位置情報の記憶)されるようですが、それがすべてを表していないようです。
シナプスの可塑性とは各々のシナプスの伝達効率が変化し、それが長時間維持されることです。そのため、学習・記憶の基本原理と考えられています。もっとも、シナプスの可塑性の研究が進んでいる分野は、海馬のNMDA受容体依存性のLTPのようです。
発現メカニズムがよくわかっている小脳のLTDは、小脳の機能への関与がまだ明確になっていないようです。
LTPの発生機序のアニメ
通常、シナプス前線維を電気刺激すると、神経終末からグルタミン酸が放出され、シナプス後膜のAMPA型受容体に結合し、受容体チャネルが開いてNa+が細胞内に流入します。
?グルタミン酸は、NMDA型受容体にも結合しますが、静止状態では、Mg+でブロックされているため、この受容体は開きません。
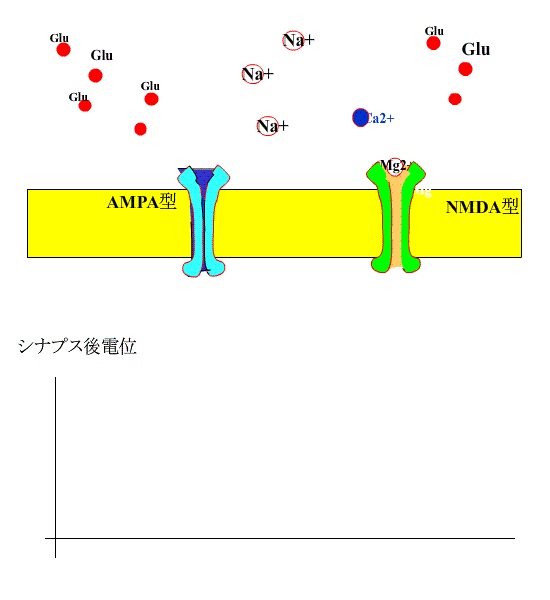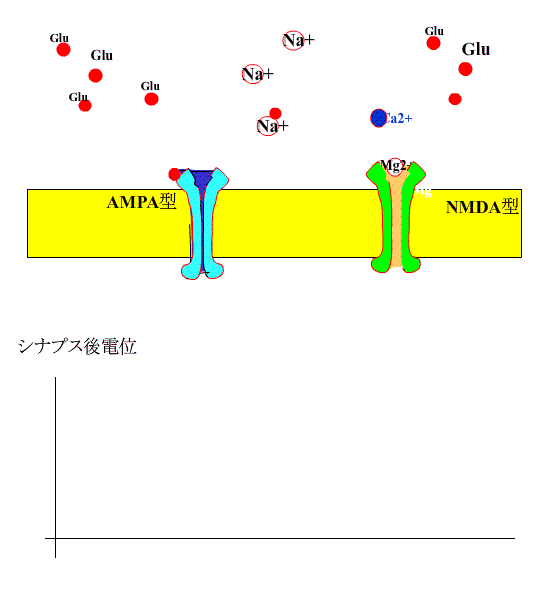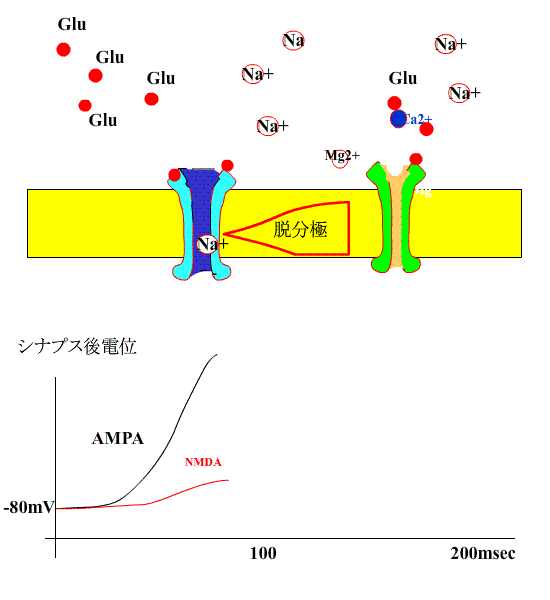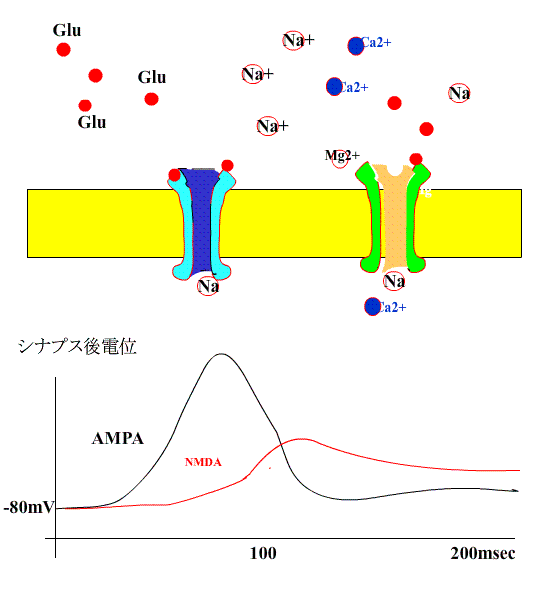 次に、シナプス前繊維を高頻度刺激すると、AMPA型受容体の活性化により、シナプス後膜が大きく脱分極され、NMDA受容体のMg+のブロックがはずれます。
次に、シナプス前繊維を高頻度刺激すると、AMPA型受容体の活性化により、シナプス後膜が大きく脱分極され、NMDA受容体のMg+のブロックがはずれます。
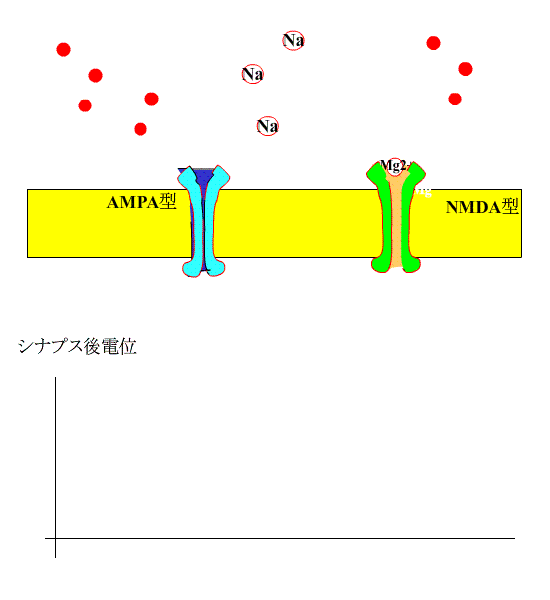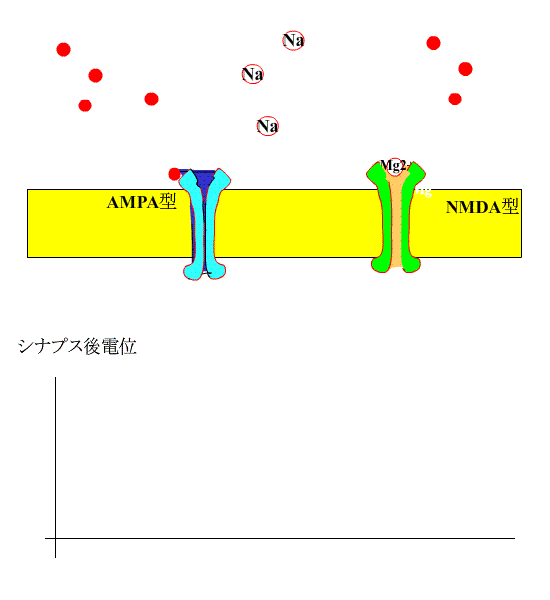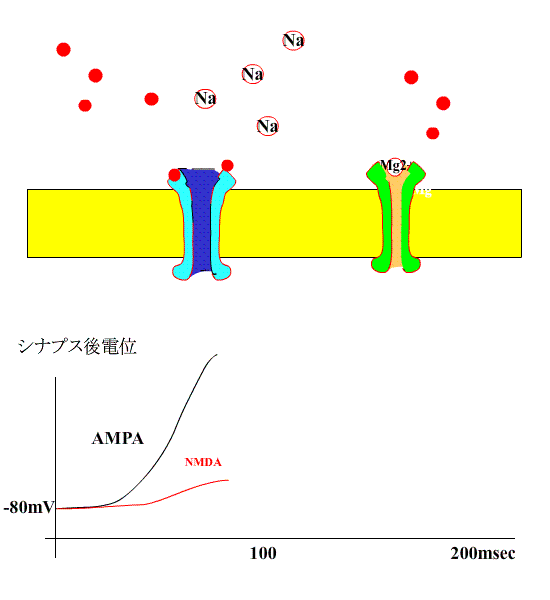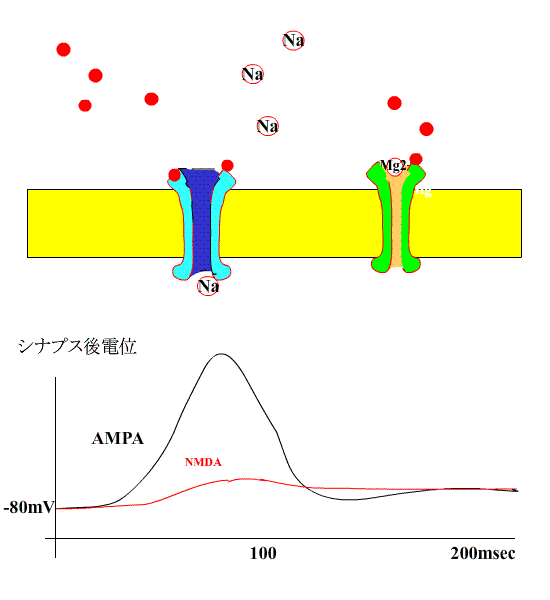
すると、NMDA型受容体のチャネルも開き、Na+に付け加えて、Ca++が細胞内に流入します。
細胞内Ca++濃度が上昇すると、Ca++依存性カルモジュリンキナーゼが活性化し、受容体の感受性が誘導されます。
このような現象が長期増強のはじめの数時間に起きているようです。
注釈)シナプス後電位とは
シナプス後電位とは何かと説明する前に、復習ですが、中枢神経系のシナプスの伝達の特徴についてです。
神経筋接合部のシナプスとは異なり、3つの特徴があります。
①1つのニューロンに対し、多数のニューロンがシナプス結合し信号を送っています。それぞれのニューロンからの単発な入力では、シナプス後ニューロンに活動電位を発生させるには、不十分です。多数の入力を統合する必要があります。
②中枢神経の神経伝達物質は多数あり、それぞれの伝達物質に対し複数の種類の受容体があります。つまり、同じ神経伝達物質でも、受容体の種類が違えば異なる応答を示します。
③興奮性と抑制性のシナプス伝達が起きることです。興奮性シナプス伝達では、神経筋接合部の伝達のように、シナプス後ニューロンに脱分極が生じます。興奮性シナプス後電位 (excitatory postsynaptic potential;EPSP )です。
一方、過分極 (抑制性シナプス後電位 inhbitory postsynaptic potential;IPSP) が生じると、活動電位の発生が抑制されます。
以上の特徴から、中枢神経系の単一ニューロンは、興奮性や抑制性の多数のシナプス入力を統合し、シナプス後電位という形で刺激を伝達します。
| AMPA受容体の増加はLTPになり、受容体の減少はLTDに |
さて、今まで、LTPとLTDはそれぞれ個別に説明が試みられてきたようですが、最近のデータでは、下のアニメにあるように、AMPA受容体の増減が、LTP/LTDを解釈するための根拠になっています。
機序を超簡単なアニメで、示します。
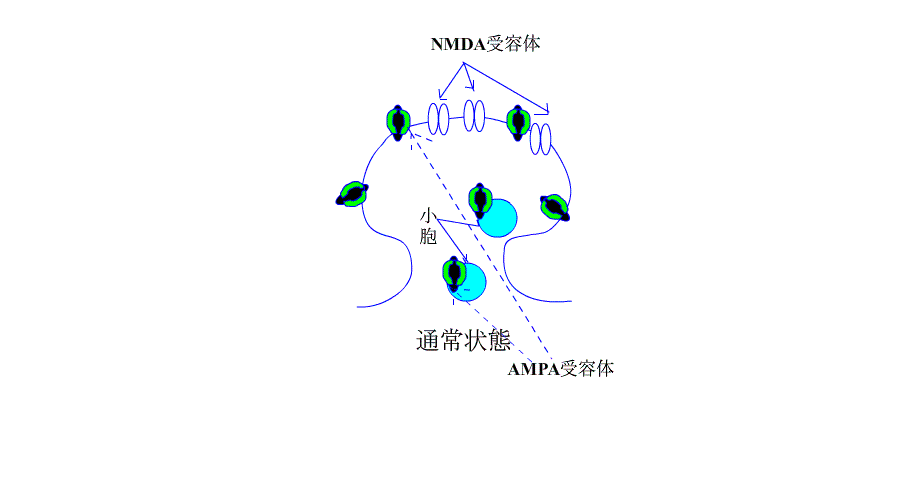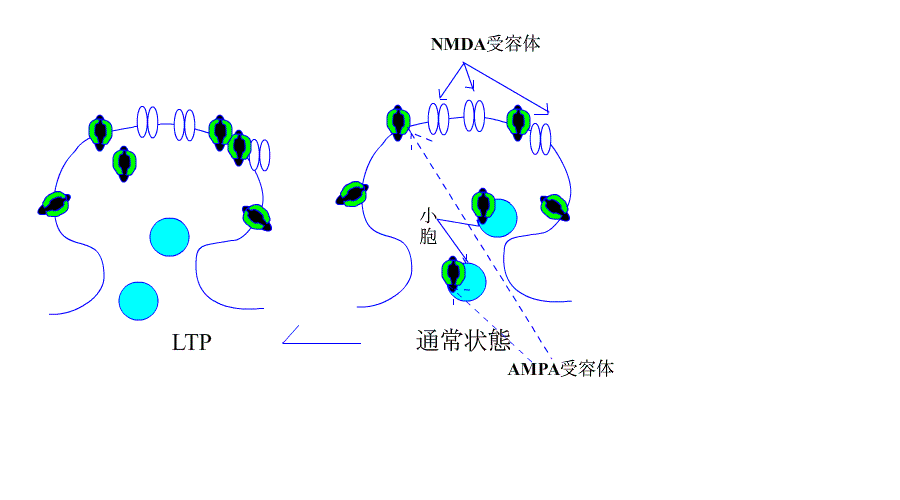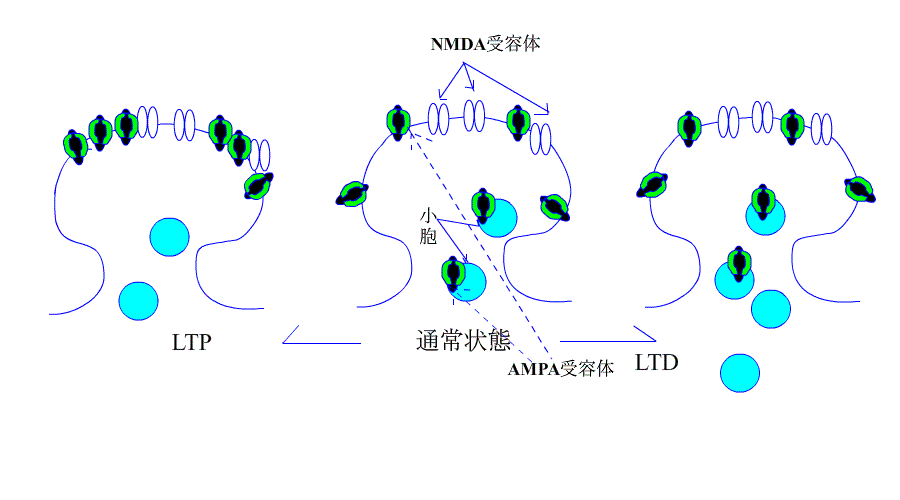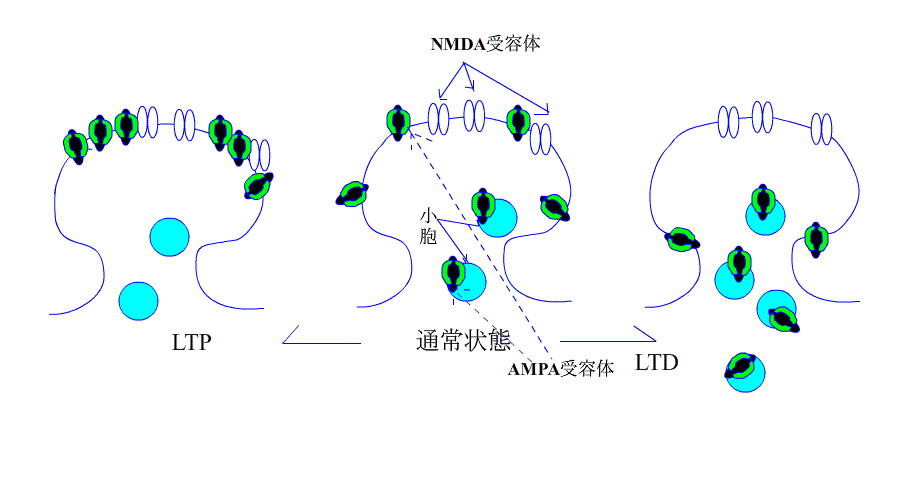
引用元:高宮考悟.学習・記憶におけるシナプス可塑性の分子貴構 生化学 第83巻 第11号.pp 1016-1026,2011
次はLTDについてです。
運動に関係する記憶(例えば自転車に乗れるようになったり、ピアノが弾けるようになったりするような記憶)の形成には、小脳のプルキンエ細胞とそこへ入力する平行線維(顆粒細胞の軸索)との間のシナプスにおいて、
神経伝達効率が長期にわたって低下することが極めて重要な役割を果たします。これを実験モデルで考えた場合、このシナプス可塑性を長期抑圧(long-term depression; LTD)と呼びます。 |
小脳について、考えてみましょう。
古くから、小脳は運動や姿勢の制御に関わると考えられてきました。
小脳皮質は、どの部分も一様で、きわめて規則的な微細構造が見られます。
巨大なプルキンエ細胞が皮質表面に平行に一列に並ぶプルキンエ細胞層
その表層の分子層
深部の顆粒細胞層の3層に区分されます。
分子層には、星状細胞とバスケット細胞が存在し、プルキンエ細胞の樹状突起が扇状に広がっています。
顆粒細胞層には、ゴルジ細胞と顆粒細胞が存在します。顆粒細胞の軸索は、皮質表面に向かって伸び、表面に至って分岐し、表面と平行に走行する平行繊維となって長く伸びます。プルキンエ細胞の樹状突起は、多数の平行繊維につき抜かれます。
5種類の細胞のうち、興奮性ニューロンは顆粒細胞だけです。他は、すべて抑制性ニューロンです。小脳皮質への入力は、苔状線維と登上繊維です。苔状線維は、脊髄、前庭神経核、橋核、脳幹網様体などから起こり、登上線維は、延髄の下オリーブ核から起こります。特に、プルキンエ細胞の樹状突起に終止し、強い興奮作用を示します。
ゴルジ細胞、バスケット細胞、星状細胞は抑制介在ニューロンです。ゴルジ細胞は、顆粒細胞からの入力により興奮し、顆粒細胞を抑制します。顆粒細胞が興奮しすぎないように負のフィードバックをかけています。
バスケット細胞と星状細胞は、平行繊維からの入力で興奮し、プルキンエ細胞を抑制します。
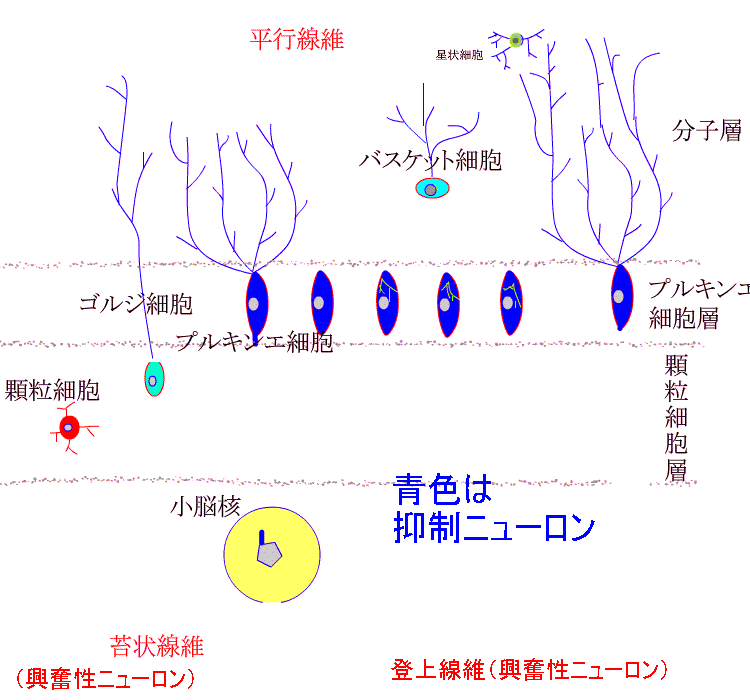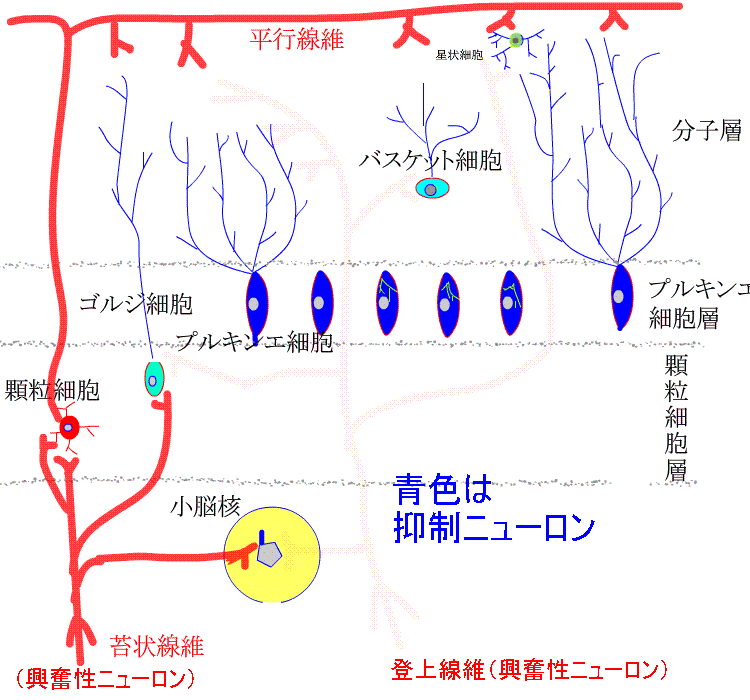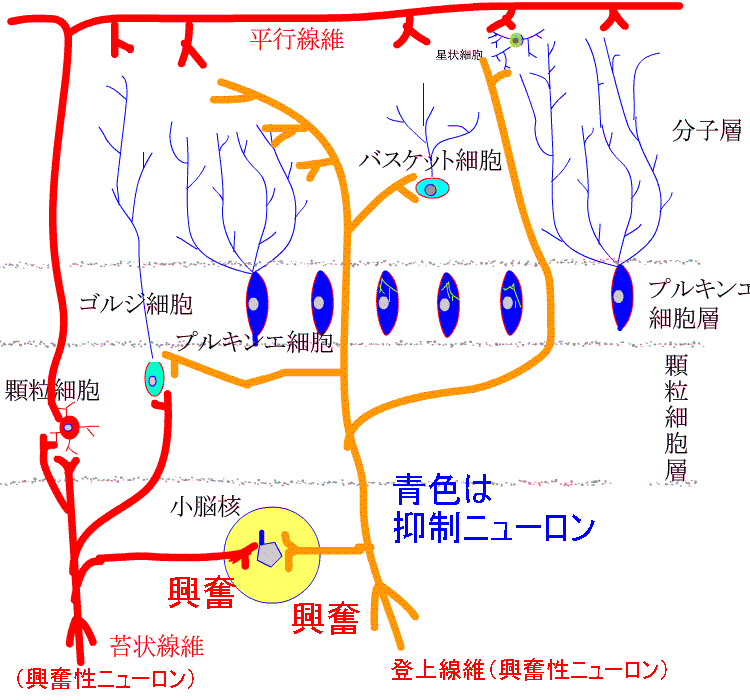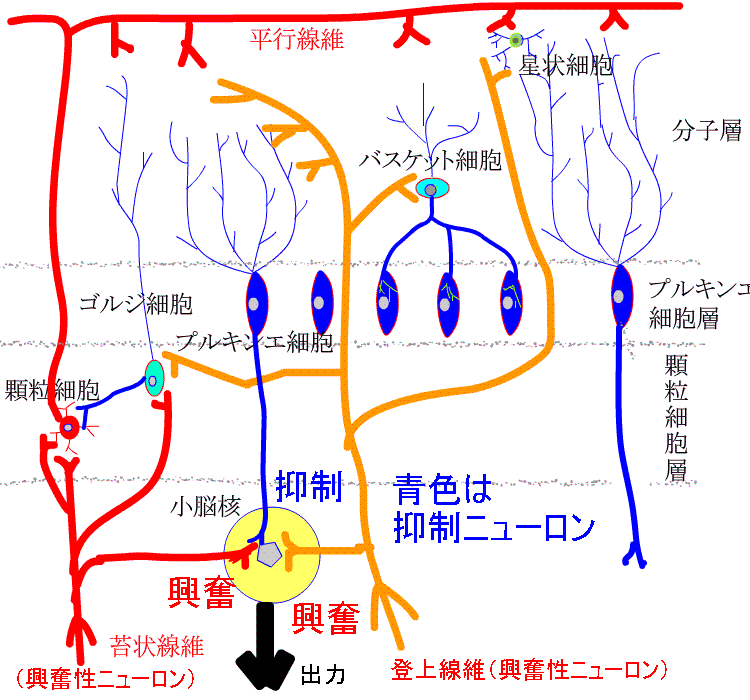
小脳皮質からの唯一の出力はプルキンエ細胞の軸索で、小脳核や前庭神経核に終止し、抑制的に作用します。
プルキンエ細胞で起こること
2系統の興奮性入力を受けます。
苔状線維から顆粒細胞そして平行線維へと伝わる入力と登上線維からの入力です。
1個のプルキンエ細胞は1万個以上の顆粒細胞と1本の登上線維から入力を受けます。
この2つの入力をうけることで、2種類の活動電位が発生します。
1つは単純スパイクで、1秒間に50~100回の高頻度で発生し、他のニューロンの活動電位と同様の形状です。平行線維からの入力で生じます。
もう一つは、複雑スパイクです。通常の活動電位と異なる多峰性の幅広い活動電位で、1秒間に約1回と低頻度で発生します。登上線維からの強力な興奮性入力で生じます。
プルキンエ細胞で、複雑スパイクが発生しているとき、平行線維から単純スパイクが繰り返し入ることで、その後、その平行線維からのシナプス伝達が長期間にわたって抑制されます。言い換えると、平行線維の活性化の2~300ミリ秒以内に登上線維が活性化した場合に長期抑圧が起こりやすい そうです。すなわち、複雑スパイクは、顆粒細胞とプルキンエ細胞との間のシナプス伝達効率を低下させます。
これがLTDになります。このLTDという動きは、顆粒細胞とプルキンエ細胞との間のシナプス伝達効率を変化させ、プルキンエ細胞の出力を適切なものに変えていると推測されるモデルのようです。
Kinate受容体
カイニン酸受容体は海馬CA3野苔状線維などの特定のシナプスに局在する傾向があります。また、AMPA型グルタミン酸受容体やNMDA型グルタミン酸受容体は興奮性シナプス伝達に直接的に寄与するのに対し、カイニン酸受容体は神経細胞の興奮性やシナプス伝達の調節因子として機能することが多いようです。
G蛋白共役型とは、グアニンヌクレオチド結合タンパク質の略称です。GTPまたはGDPを結合して活性を発揮することで、細胞内情報伝達に関与します。
Gタンパク質は、グアニンヌクレオチドを結合しGTPアーゼ活性をもつαサブユニットとβ、γサブユニットからなる3量体のタンパク質で、Gタンパク質共役受容体(GPCR)により活性化されます。
また、αサブユニットは、GTPアーゼドメインとαヘリクッスドメインからなり、20種類以上のαサブユニットがあるようです。アデニール酸シクラーゼを活性化し、cAMPを合成し増やしたり、逆に、アデニール酸シクラーゼを抑制したりします。ほかには、ホスホリパーゼCを刺激したり、網膜のロドプシンと結合し、視覚のシグナルを伝達するようです。他にも、沢山、情報があります。
注)サブユニットとドメインについては、以下のサイトを見てください。
https://satoayumi.com の高校生物/大学生物の後半部分を見てください。分かりやすいです。
次に、セカンドメッセンジャーのcAMPを合成し増やすメカニズムを簡単なアニメで示してみます。

?
?
Gタンパク質共役受容体に結合する赤い点は、アドレナリン、グルカゴンまたは黄体形成ホルモンです。
セカンドメッセンジャーとして、cAMP(サイクリックAMP)が生成されます。このcAMPによって、タンパク質キナーゼA(A Protein KinaseA : PKA)すなわち、cAMPー依存性タンパク質キナーゼが働き、細胞質の分子活動を変化させます。このPKAは、タンパク質をリン酸化する酵素です。
次に、本題のGタンパク質共役受容体に結合するグルタミン酸の場合について考えてみます。
代謝型グルタミン酸受容体はグループ I(mGlu1および mGlu5)、グループ II(mGlu2および mGlu3)、グループ III(mGlu4、mGlu6、mGlu7および mGlu8)の 3 グループに分類されます。
グループ Ⅰ 代謝型グルタミン酸受容体は主にシナプス後膜に局在し、シグナルを受けると ホスホリパーゼC(PLC )を活性化して細胞内のジアシルグリセロール(Diacylglycerol)およびイノシトール -3- リン酸(Inositol triphosphate)の濃度を増加させます。これによりプロテインキナーゼC(PKC) が活性化し、小胞体などに貯蔵されていた Ca2+が細胞内に放出されて細胞内カルシウム濃度が上昇します。その結果シナプス前膜の K+チャネルを抑制し、神経終末端での脱分極を遅延し、シナプス伝達を抑制します。
グループ Ⅱ 代謝型グルタミン酸受容体は主にシナプス前膜に局在し、アデニル酸シクラーゼ活性を阻害することで興奮性神経伝達を抑制します。
グループ Ⅲ 代謝型グルタミン酸受容体はグループ Ⅱ と同様、主にシナプス前膜に局在し、アデニル酸シクラーゼ(Adenylate cyclase)活性を阻害することで興奮性神経伝達を抑制します。